

Botany & Plant Pathology
Planting seeds of innovation and discovery for plant sciences since 1887.
Join Botany and Plant Pathology at Purdue
CONTACT US
Department of Botany and Plant Pathology
Lilly Hall of Life Sciences, Room 1-446
915 Mitch Daniels Boulevard
West Lafayette, IN 47907
(765) 494-4615
botany@purdue.edu
New & Noteworthy
New & Noteworthy
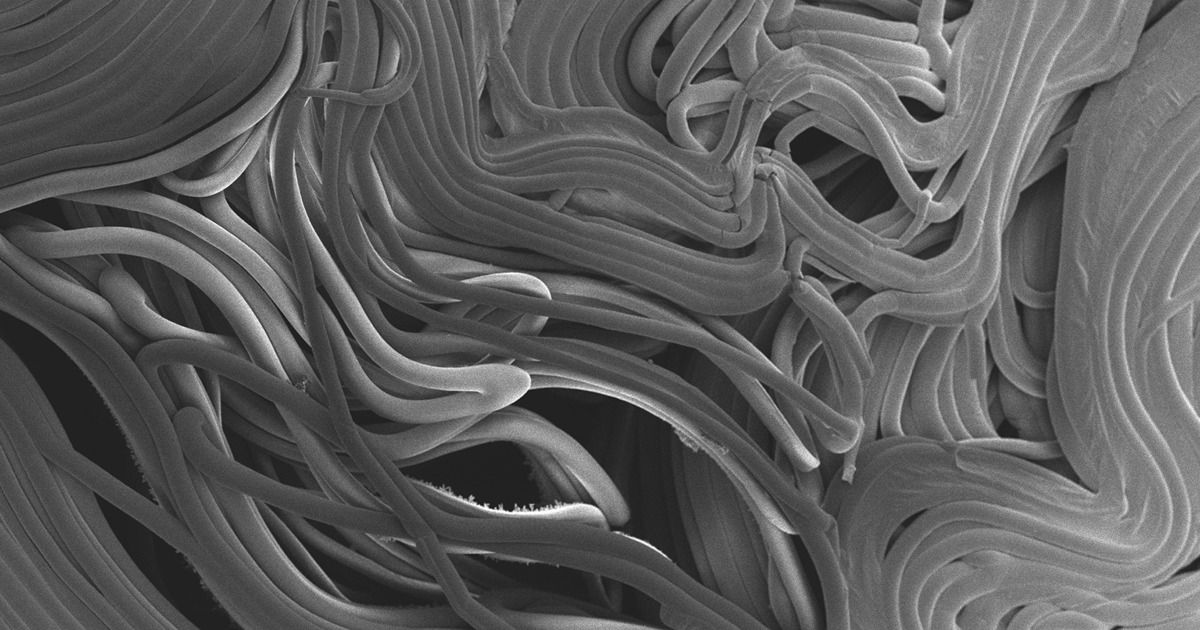
Daniel Szymanski and his former postdoctoral scholar Youngwoo Lee, lead author on their paper recently published in Plant Physiology, analyzed proteins in developing cotton fibers over time. They partnered with Jonathan Wendel, distinguished professor of ecology, evolution, and organismal biology, and Olga Zabotina, a professor of biochemistry biophysics and molecular biology, both from Iowa State University, as well as Jun Xie, a professor of statistics at Purdue, to research the multiple dimensions responsible for cells behind the billion dollar industry.

One of the final projects funded by Plant Sciences 2.0, one of Purdue’s Next Moves strategic initiatives, were the new set of high-tech greenhouses in the Lilly Life Science Ranges, which was completed in the fall of 2025. The newest 18,654 square feet of greenhouses continue Purdue’s long legacy of putting cutting-edge technology into Boilermakers’ hands.

Purdue researchers in the colleges of Agriculture, Engineering and Science are collaborating to advance AI applications.

The eighth wonder of the world might be a hundred-million-year-old process: pollination. When you stop and smell the flowers or buy a bouquet, aroma and beauty likely drew you in. Meanwhile, right under your nose, ancient partnerships are hidden within every bud and are the reason for the sweet scent, bright colors, intricate patterns and up to 90% of the food you eat. This floral complexity is not made for human appreciation, but rather to move tiny particles that make us sneeze and rub our eyes — pollen grains.

The average long-term return on the stock market is about 10 percent. For AgSEED, the Purdue College of Agriculture’s internal competitive grants system, it’s more like 400 percent. It’s an outsized return with outsized impact, benefiting researchers, students, and citizens across the state and beyond.

Camila Nicolli, research assistant professor of mycotoxin fungal biology in the Department of Plant Pathology, and Matheus Zavadinack, research assistant professor in carbohydrate chemistry in the Department of Food Science, have joined Purdue University's College of Agriculture faculty this Spring semester.
What is a Botanist?
From the smallest moss to giant sequoia trees, from lichen to mushrooms, from corn to the bacteria and fungi that grow with it — botany goes beyond the study of plants. Botanists are plant scientists, curious about the natural world and how living things grow, interact with each other and change their environment.
Botany and Plant Pathology research protects crucial ecosystems, influences conservation and natural resource management and secures humanity’s food supply with discoveries in genetics, microbiology, plant health, mycology, ecology, anatomy and weed control.
Cutting-edge science, rich in history
Formally established in 1887, Botany and Plant Pathology has a deep-rooted legacy at Purdue. One of the six original professors at Purdue University was botanist Reverend John Hussey, and the first doctoral degree given by Purdue was to Daniel T. MacDougal, a botanist with a thesis titled, “The Curvature of Roots.”
Botany and Plant Pathology is home to 32 faculty members making innovations and discoveries that span plant health, anatomy and physiology, microbiology, environment and ecology, genetics and mycology.
Botany is one of the oldest sciences. People around the world have long studied plants and fungi for food, materials and medicine. Purdue University’s One Health initiative is kickstarting a new age of interdisciplinary research connecting plant science to environmental, human and animal health. Plant science research is more important today than ever before to feed and care for the growing global population and protect the Earth that supports it.
Can I be a botanist?
Are you interested in how plants and fungi grow? Do you wonder about the complex relationships between plants, microbes and their environment? Want to know how cells change to stretch their leaves and stems toward the sun? You may be a botanist in the making — join Botany and Plant Pathology’s plant science major!
"Purdue University is a hub for plant science research, with diverse flavors of fundamental science that enriches knowledge of plants, to how they grow, to translating that knowledge to solve current and emerging societal challenges. To keep up with global population growth and a changing planet, we are bringing botany into the 21st century with a tight-knit but interdisciplinary community of scientists and students."
Tesfaye Mengiste
Botany and Plant Pathology Department Head
Tesfaye Mengiste Botany and Plant Pathology Department Head
"Botany and Plant Pathology at Purdue houses the world’s most important collection of rust fungi — obligate plant pathogens that can cause devastating disease in agricultural crops and natural ecosystems. Purdue has been a global center for the study of these fungi for more than 100 years."
M. Catherine (Cathie) Aime
Professor of Mycology
M. Catherine (Cathie) Aime Professor of Mycology
"I love the hands-on lab experiences that make plant biology come to life and the excitement that students bring to discovering how plants shape our planet. The enthusiasm in botany is contagious."
Morgan Furze
Assistant Professor of Botany and Forestry
Morgan Furze Assistant Professor of Botany and Forestry

"The botany department at Purdue University is a wonderful place to integrate both basic and applied plant research. Our comprehensive curriculum and collaborative research environment provide students with outstanding opportunities to engage in hands-on research, work closely with experts across disciplines, and prepare for a wide range of career paths in plant science."
Yun Zhou
Associate Professor of Plant Developmental Biology
Yun Zhou Associate Professor of Plant Developmental Biology
Subscribe & Support
Botany and Plant Pathology is a community that includes you. Our Extension agents, farmers and the people they feed are crucial parts of our research. Subscribe to our newsletter, Root of the Matter, to stay updated on our latest discoveries and impacts around the world. Your support and donations help us continue to push the boundaries of plant science and beyond.
Facilities & Communities
Scientists need more than just a lab to flourish. The College of Agriculture and the Botany and
Plant Pathology Department connects faculty, staff and students to a variety of facilities and
communities on campus to help them, their research and their outreach grow.





